The taxonomy of myomorph rodents has undergone various perturbations over the last few decades. In earlier times, the Old World rats and mice were placed in the family Muridae and the New World rats and mice in the family Cricetidae (neither of the common names for the groups are especially appropriate). Later, all were placed into the Muridae. Currently, the classification is back to considering there to be two families, but with the subdivisions named differently than in the early classification. To be able to use the literature, it is necessary to understand the changes that have occurred.
In the older tradition, the families Muridae and Cricetidae were separate, with the Cricetidae divided (for our region) into the Cricetinae and the Microtinae. More recently, all were considered to fall within the Muridae, with our representatives divided among the subfamilies Murinae (Old World mice and rats), Sigmodontinae (mice and rats with generally non-prismatic teeth; not vole-like), and Arvicolinae (mice and rats with prismatic teeth; vole-like; equal to the earlier Microtinae). Most recently (Wilson and Reeder, 2005), the Muridae and Cricetidae have been recognized once more as separate families; however, the Cricetidae has been recognized as consisting (within our region) of three subfamilies: those earlier recognized as sigmodontines have been split into two subfamilies, the Neotominae (Woodrat and peromyscine types) and the Sigmodontinae (forms with southern affinities). The third subfamily remains the Arvicolinae (voles, sensu lato).
The Muridae is a huge family, but most are Old World. There are only three New World representatives, all introduced: the Norway Rat (Rattus norvegicus), the Black Rat (Rattus rattus), and the House Mouse (Mus musculus). These will be considered in the Muridae module.
We will be considering only the small portion of this family that occurs in our region of interest. In such a large family, a wide diversity of morphological, ecological, and behavioral traits is expected—and found. Characteristics in common beyond the myomorphus condition include a reduced dental battery of formula 1/1 0/0 0/0 3/3 = 16 (with a few exceptions). The complexity of the cheek teeth varies greatly, but the basic pattern is of five crests separated by reentrants.
Subfamily Neotominae and Subfamily Sigmodontinae—New World Rats and Mice
The Neotominae is a large subfamily that includes most of the mice and rats that we are familiar with. Most neotomines are more or less adapted to a generalized terrestrial or climbing (scansorial) life style, but some arboreal, semi-aquatic, and fossorial taxa are included. The limb structure tends to be relatively generalized, with fusion (when it occurs) generally limited to the tibia and fibula.
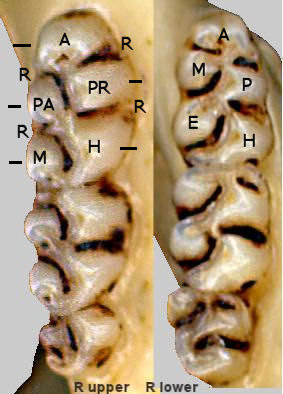
Fig. 1. Upper and lower toothrows of Peromyscus showing crests (cusps) and reentrants ("R"). Short bars indicate the five crests. The overall aspect is typical of the "peromyscine" genera. A = Anterocone (upper), Anteroconid (lower); PR = protocone (upper), P = protoconid (lower); PA = Paracone; M = Metacone (upper), Metaconid (lower); H = Hypocone (upper), Hypoconid (lower); E = Entoconid.
The local forms generally are large-eyed, large eared, and long-tailed forms that are primarily crepuscular and nocturnal. There are, however, exceptions, with the grasshopper mice being short-tailed. The only genus of the Sigmodontinae within our region (Sigmodon; cotton rats) often are active diurnally. Most members of the two subfamilies are relatively sharply bicolor, the undersides distinctly lighter than the dorsal surfaces and the transition usually relatively sharp.
Regional taxa are listed in the classification section. The genera Peromyscus and Neotoma are ubiquitous in our area and generally have well defined niches. The remaining genera are less than universal in distribution, and one (Baiomys) is geographically marginal, entering the New Mexico/Trans-Pecos Texas region only in the bootheel of southwestern New Mexico.
At the generic level, these rodents are rather easily distinguished from one another. Neotoma has hypsodont, semi-prismatic cheek teeth; Sigmodon has brachyodont teeth with rather complex (but not prismatic—see Sigmodontinae/Neotominae 3) occlusal patterns; the remaining genera have brachyodont, more or less bunodont, dentition. Among these, the harvest mice (Reithrodontomys) are the only group to have grooved upper incisors. The grasshopper mice (Onychomys) have a short tail and molars that have widely open reentrants without stylets or cusplets within the reentrants. Baiomys can be told from Peromyscus primarily on the basis of size: head and body length generally is less than 65 mm, hind foot length less than 20 mm, and occipitonasal length less than 22 mm; the skin of Baiomys is more apt to be confused with that of the House Mouse (Mus musculus) than that of Peromyscus.
Information on a number of Southwestern rodents can be found on the Sevilleta site. The Animal Diversity Web is a great resource for rodents, as well as other groups. The Smithsonian Field Guide series also is very useful for the neotomines, sigmodontines, and arvicolines.
Baiomys taylori (Pygmy Mouse) occurs in dense grass in Hidalgo County, southwestern New Mexico, apparently constructing runways and perhaps with more diurnal activity than seen in the deer mice. It appears to feed more extensively on the vegetative portions of plants than Peromyscus. Another difference from the deer mice is in parental care, the males seemingly aiding in the care of the young.
In captivity, Pygmy Mice produce average litters of three at about 1-month intervals. According to Findley (1987), the young appear to remain fastened to the nipples of the female until weaned at 18 to 22 days.
 Neotoma—with six species in the region,
Woodrats (also known as packrats) are common in most habitats.
Neotoma—with six species in the region,
Woodrats (also known as packrats) are common in most habitats.
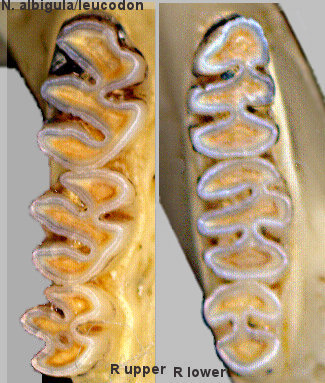
Fig. 2. Upper and lower cheek-tooth rows of a White-throated Woodrat. The teeth are typical of the genus Neotoma.
One of their most interesting habits is that of constructing middens of plant and other material, either surrounding the nest or, in protected situations, sometimes also nearby. The middens have been of great help to workers attempting to reconstruct the nature of the vegetation in past times. Woodrats limit their home range to within about 100 m of the nest and thus give a good picture of the local vegetation at any one time. Since the middens are of organic material, they are easily dated by 14C dating, and middens protected in caves and dry rock crevices have been dated back to about 40,000 radiocarbon years BP (before present). Urination onto sheltered middens may result in the formation of amberat (the dried, fossil urine) which appears to have some antibacterial effect, aiding in the preservation of plant material.
Middens often are occupied for many generations of Woodrats and thus may record vegetative changes through time, with the various layers being individually datable. Middens, occupied or abandoned, often are utilized by other animals, including other rodents, reptiles, shrews, lagomorphs, and invertebrates. The remains of these, plus "pickups" by the Woodrats, may give further environmental information. A wealth of information on modern and fossil Woodrat middens is available in Betancourt et al. (1990).
 Woodrats generally are antisocial except when the female is in
estrus; thus usually only one adult rat occupies and defends the nest. Nests in favorable positions usually are taken over by other Woodrats if the owner dies. Breeding is seasonal, though the
season varies with species and elevation. In the lowlands, it may begin in January; at high elevations, it may not end until August. Although one to six offspring may make up a litter, the average
number is toward the low end of this range. Weaning is at 4 to 6 weeks, and adult size is attained by about the 8th month; nevertheless, some of the lowland, smaller species may begin to breed by 2
to 3 months of age. The high-elevation Bushy-tailed Woodrat, however, doesn't breed until its 2nd year.
Woodrats generally are antisocial except when the female is in
estrus; thus usually only one adult rat occupies and defends the nest. Nests in favorable positions usually are taken over by other Woodrats if the owner dies. Breeding is seasonal, though the
season varies with species and elevation. In the lowlands, it may begin in January; at high elevations, it may not end until August. Although one to six offspring may make up a litter, the average
number is toward the low end of this range. Weaning is at 4 to 6 weeks, and adult size is attained by about the 8th month; nevertheless, some of the lowland, smaller species may begin to breed by 2
to 3 months of age. The high-elevation Bushy-tailed Woodrat, however, doesn't breed until its 2nd year.
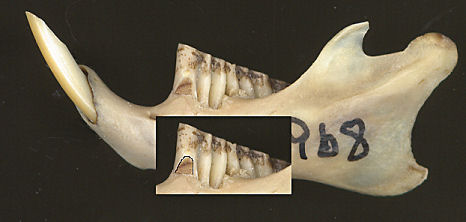
Fig 3. Left dentary of a woodrat (Neotoma). The inset has the lateral dentine tract on m1 delimited by the black line.
Characters separating the various species include the development of the lateral dentine tract on m1 (Fig. 3). The species usually occurring woodland or higher-elevation habitats (N. cinerea, N. mexicana, N. stephensi) have the tract better developed than the lowland species (N. albigula, N. leucodon, N. micropus) (Harris 1984).
Woodrats feed most extensively on the foliage of plants, though other material, including invertebrates, will be eaten.
 Neotoma albigula (White-throated
Woodrat) is the common Woodrat in the southern lowlands of New Mexico west of the Rio Grande. East of the Rio Grande and throughout Trans-Pecos Texas, the White-toothed Woodrat (Neotoma
leucodon) replaces it. The latter is indistinguishable from the former on morphological characters (establishment as a separate species is on the basis of DNA data). Members of the pair of
species are common from southern Colorado south into central Mexico. They inhabit grassland, desert, and pinyon-juniper woodland. In favorable areas, density reaches some 10 per acre, and over 20
per acre have been recorded. Where sympatric with the Southern Plains Woodrat (N. micropus) in eastern, southern, and southwestern New Mexico and in Trans-Pecos Texas, they tend to stick to
rocky areas, though occasionally venturing out into desert basins and building nests within mesquite clumps or similar protective vegetation. Where N. micropus is absent, they spread into
broader habitats.
Neotoma albigula (White-throated
Woodrat) is the common Woodrat in the southern lowlands of New Mexico west of the Rio Grande. East of the Rio Grande and throughout Trans-Pecos Texas, the White-toothed Woodrat (Neotoma
leucodon) replaces it. The latter is indistinguishable from the former on morphological characters (establishment as a separate species is on the basis of DNA data). Members of the pair of
species are common from southern Colorado south into central Mexico. They inhabit grassland, desert, and pinyon-juniper woodland. In favorable areas, density reaches some 10 per acre, and over 20
per acre have been recorded. Where sympatric with the Southern Plains Woodrat (N. micropus) in eastern, southern, and southwestern New Mexico and in Trans-Pecos Texas, they tend to stick to
rocky areas, though occasionally venturing out into desert basins and building nests within mesquite clumps or similar protective vegetation. Where N. micropus is absent, they spread into
broader habitats.
These Woodrats are perfectly at home with cacti and clamber over them with seeming impunity, feeding on the fruits and pads of prickly pear and with the nest often strongly defended by joints of prickly pear and cholla and, more painfully, by the spiny areoles left after feeding on the pads.
Most cave fossil sites in our region produce remains of the White-throated Woodrats pretty much through the late Pleistocene record.
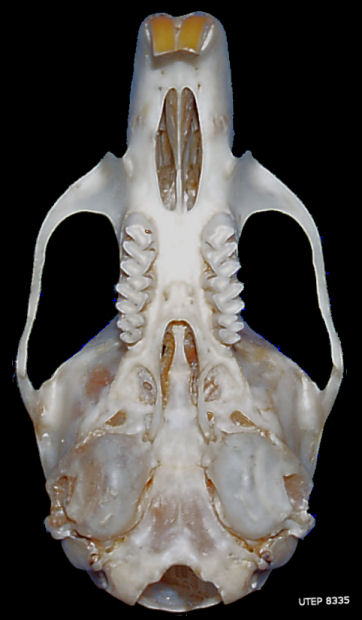
Fig. 4. Ventral view of the skull of the White-throated Woodrat (Neotoma albigula). The general overall character of the skull is common to all of the species.
Last Update: 12 Feb 2008
Centennial Museum and Department of Biological Sciences, The University of Texas at El Paso