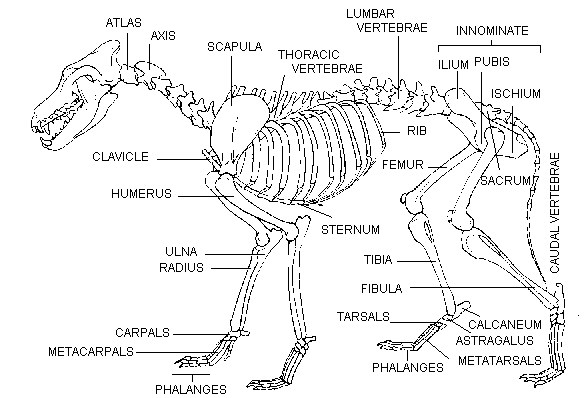
A wolf skeleton showing most of the major skeletal elements. The atlas, axis, and the (almost always) five next vertebrae make up the cervical vertebrae.
Bone is a combination of mineral and organic materials. The mineral material gives strength and the organic matrix prevents 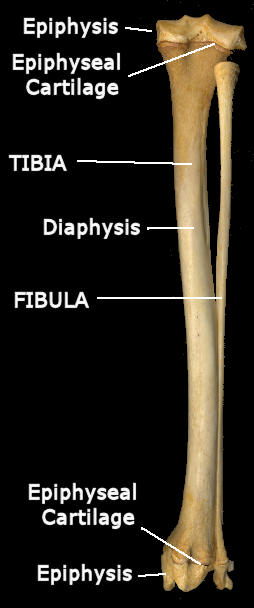 the extreme brittleness that mineral alone would provide. Although we tend to think of bone as an inert material, it is far
from that. Among other things, the skeleton acts as a store of calcium and phosphorus, releasing or taking up these elements as homeostasis requires. Bone also is constantly being remodeled and
will actively react to stresses. Heavy muscle use, for example, will cause enlargement of tendon attachment areas (one of the reasons for the more gracile human female skeleton compared to that of
the male). Much of the remodeling of bone is done by internal reworking.
the extreme brittleness that mineral alone would provide. Although we tend to think of bone as an inert material, it is far
from that. Among other things, the skeleton acts as a store of calcium and phosphorus, releasing or taking up these elements as homeostasis requires. Bone also is constantly being remodeled and
will actively react to stresses. Heavy muscle use, for example, will cause enlargement of tendon attachment areas (one of the reasons for the more gracile human female skeleton compared to that of
the male). Much of the remodeling of bone is done by internal reworking.
Most of the deeper bones of the body (and some more surficial) start out as cartilage in the embryo (and thus, amazingly enough, often are classified as "cartilage bone" in contrast to "membrane or dermal bone" which forms without a cartilage predecessor). Most of the mammalian long bones such as the limb bones, along with vertebrae, turn into bone (ossify) from more than one center of ossification. Thus in an idealized long bone, there would be three such centers (some bones have more centers): one at each end and one toward the center of the shaft. The main shaft is the diaphysis; the ossifications at the ends are epiphyses. The growing centers of the diaphysis and each epiphysis grow toward one another, though the diaphysis lengthens considerably faster than the epiphyses. Eventually, all will meet and fuse into a single ossified structure. In general, however, such fusion occurs long after birth and different epiphyses fuse at different times, allowing the possibility of aging a mammal by the pattern of epiphyseal fusion. Before such fusion occurs, the epiphyses are separated from the diaphysis by a plate of cartilage, the epiphyseal cartilage. During the fusion process, there often is a discernable epiphyseal line left in the bone. (Diaphysis bone near the epiphysis in a young animal usually has a distinctive texture unlike the hard-surfaced bone of an adult; this is another sign of an immature animal.)
Posterior view of the right tibia and fibula of a young Red Fox (Vulpes vulpes) showing epiphyses and diaphyses. Fusion is nearly complete, so relatively little epiphyseal cartilage is left.
Cartilage has an advantage over bone in that it can enlarge by internal growth, whereas bone can enlarge only by accretion to the surface. Having epiphyses presumably provides a stable articular surface at joints while the epiphyseal cartilage allows for continued growth in length. Once fusion has occurred, further growth is, for all practical purposes, complete.
The common ancestor of mammals presumably had five digits on each limb, and such limb bones as radius and ulna, and tibia and fibula were completely separate elements. During the course of evolution, there has been a variety of losses, fusions, and a few additions in various taxa. Frequently, the tibia and fibula have partially fused into a tibiofibula; less commonly, the ulna and radius have fused. Loss of digits is widespread, even to the point where only a single digit per foot survives in the horses. Some of the various skeletal differences will be noted in the accounts of taxa.
Last Update: 9 Jan 2009
Centennial Museum and Department of Biological Sciences, The University of Texas at El Paso