Geologic time (or that portion we need to be familiar with) is divided into a hierarchy consisting of eras which consist of two or more periods which consist of two or more epochs. Epochs may be divided into ages. The fossil record of the vertebrates falls into three geologic eras: the Paleozoic, the Mesozoic, and the Cenozoic. The figure below on the left (courtesy of the Geologic Time Scale Foundation) shows these eras. Ages are given in millions of years. The Neogene and Paleogene only recently have been elevated to periods, replacing the Tertiary Period, which you will find in most of the older literature; most of the older literature also will give the beginning of the Pleistocene as 1.8 million years ago, a figure that has been superseded.
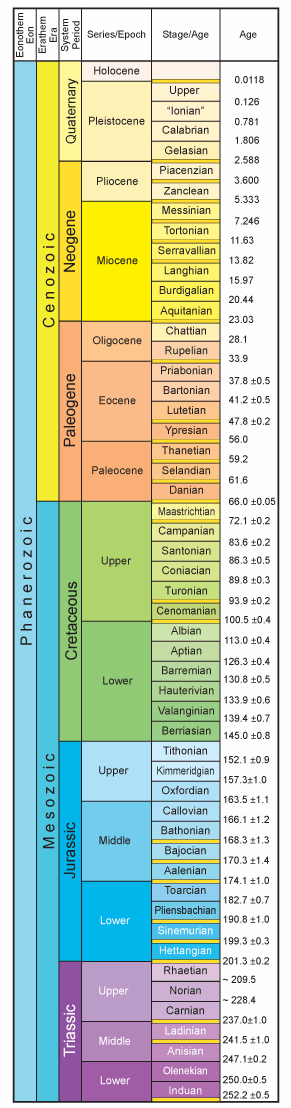
The Cenozoic starts at about 66 mya: the K-P boundary (Cretaceous-Paleogene boundary, formerly the K-T [Cretaceous-Tertiary] boundary). A major extinction event took place at the boundary. At that time, the dinosaurs, among many other groups, such as ammonites and pterosaurs, became extinct. Traditionally, it is assumed that this extinction left open ecological niches that were taken advantage of by the mammals, which promptly radiated into numerous niches to become the dominant, terrestrial vertebrate lifeform (according to the mammals, who write the histories—birds would write it differently). Recent research on the time of origin of the various mammalian phylogenetic lines, however, suggest that most major lines originated well back in the Mesozoic. It should be noted that some two-thirds of the history of mammals occurred previous to this time, with relatively modest diversity.
Mammals evolved from reptiles (using classical terminology) during the Mesozoic. The group of reptiles giving rise to the mammals were members of the subclass Synapsida. Primitively, land vertebrates had a braincase of cartilage bone and an external shield, or skull roof, of dermal bone. In several lines of reptiles, one or more openings developed in the skull roof, apparently to allow for more efficient jaw musculature. In the synapsids, the opening was relatively low on the side of the skull, with the postorbital (later lost evolutionarily) and the squamosal bones making contact above the opening. The space between the skull roof and the braincase is taken up mostly by muscles associated with movements of the lower jaw.
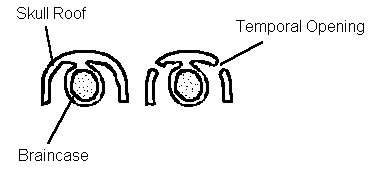
Diagrammatic cross section showing the relationship between skull roof, braincase, and temporal opening.
Two orders of synapsids evolved sequentially, with the Therapsida evolving from the earlier Pelycosauria. Therapsids often are called mammal-like reptiles since they evolved characters typical of mammals and one or more lines gave rise to the mammals. One of the suborders of pelycosaurs was the Sphenacodontia, the dominant carnivores during the earlier part of the Permian (the last period of the Mesozoic). The sphenacodonts are associated together by a feature of the lower jaw: the angular bone at the posterior ventral portion of the jaw had a reflected lamina (that is, a flange of bone flaring out to the side) that helped support the tympanum (eardrum). The sphenacodonts eventually gave rise to the therapsid synapsids in the middle or late Permian.
Therapsids evolved both herbivorous and carnivorous lines. The latter, the subclass Theriodontia, in turn radiated into several groups, most of which showed at least some of the mammalian characteristics. One group, the Cynodontia, approached the mammalian grade in several lines. Characters seen include development of a secondary palate, differentiation of cheek teeth into premolars and molars, gain of three cusps on the molars (tricuspid), double roots for the cheek teeth, increase in size of the dentary bone along with reduction of the post-dentary bones, contact of the squamosal and dentary, attainment of the mammalian phalangeal formula (number of phalanges in the digits, which primitively are 2, 3, 3, 3, 3; the formula that humans retain), appearance of the atlas and axis as differentiated cervical vertebrae, modification of the pectoral and pelvic girdle toward the mammalian condition, and further differentiation of the vertebrae.
The transition to mammals was so continuous that the separation between the two classes is arbitrary. Various schemes have been used, but probably the simplest (if not universally accepted) is drawing the line at the point where the dentary articulates with the squamosal and the quadrate and articular no longer are part of the jaw joint (an intermediate condition is known where the joint is formed by all four bones, the dentary and squamosal laterally and the quadrate and articular medially). Various forms generally placed as mammals retained post-dentary elements, but these were small and only loosely attached.
The transition point between therapsid and mammal generally is considered to be late Triassic (with therapsids continuing on into the middle portion of the Jurassic before becoming extinct). Most mammalogists consider the Mammalia to be monophyletic, but polyphyly is possible. The question hinges upon whether the lines leading to the Prototheria (represented today by the egg-laying mammals) and the remaining mammalian groups split shortly before achieving full mammalian status and evolved the mammalian grade in parallel or whether the split was after the mammalian grade was achieved.
Early Mesozoic mammalian evolution is poorly known. There obviously were several lines evolving, but the relationships among them are only dimly understood. There also is some disagreement among workers as to whether some forms should be considered mammals or as extremely closely related synapsid reptiles. Among forms considered by many to be mammals but not so considered by others are the families Sinoconodontidae, Haramiyoidea, Docodontidae, and Morganucodontidae.
The infraclass Triconodonta (late Triassic to late Cretaceous) is considered as mammalian by virtually everyone. The name comes from the molars, which have three cusps arranged in a single file, generally with the middle cusp being the best developed.
Within the infraclass Holotheria, the superlegion Kuehneotheria is known from the late Triassic to early Jurassic of Europe. Members of the taxon, such as Kuehneotherium, are characterized by the possession of tribosphenic teeth. The molars of modern mammals apparently have been derived from tribosphenic teeth.
One other major group of fairly early mammals was the Multituberculata. This sole order of the infraclass Allotheria arose in the middle Jurassic and survived through most of the Eocene, apparently until outcompeted by the emerging Rodentia. During this 120+ million years, they were a successful herbivorous (mostly) group, ranging from small mouse size to about the size of a woodchuck. A single pair of lower incisors and up to three pair of upper incisors were typical, but the most characteristic feature was the possession of many cusped molars (and thus the name Multituberculata). Some taxa also had a spectacular bladelike fourth premolar.
Althogether, mammals remained small and probably nocturnal animals throughout the Mesozoic. The generally given reason for the perceived lack of progress is competition with the dinosaurs, but although the lack of overlap in size between the taxa probably is significant, direct competition seems otherwise unlikely. In the last few years, new fossil material indicates Mesozoic mammalian diversity was considerably greater than earlier appreciated. For example, forms adapted for gliding and for swimming are now known.
Last Update: 11 Jul 2014: updated geological timescale
Centennial Museum and Department of Biological Sciences, The University of Texas at El Paso